کیا سم کارخانه تولید کود کشاورزی | کود کشاورزی ، سم کشاورزی
کیا سم کارخانه تولید کود کشاورزی | کود کشاورزی ، سم کشاورزی

امروزه گسترش کشاورزی به اندازه ای است که نمیتوان آن را در مقیاس منطقه ای بررسی کرد و باید به صورت جهانی به آن نگریست. با توجه به اینکه زندگی روی کره زمین وابسته به انرژی خورشید بوده و فتوسنتز به عنوان تنها فرآیند مهم بیولوژیکی قادر به بهره برداری از این انرژی می باشد، لذا باید با شناخت ویژگی های مسیر فتوسنتزی گیاهان مختلف و درک روابط بین گیاه و محیط به حداکثر توان برداشت از این رابطه اقدام نمود. فتوسنتز جذب انرژی خورشید به وسیله گیاهان سبز را میسر می کند، بنابراین میتوان این فرآیند را یک مکانیسم انرژی خواه دانست که انرژی جذب شده را در مولکول های بزرگ آلی ذخیره می سازد.
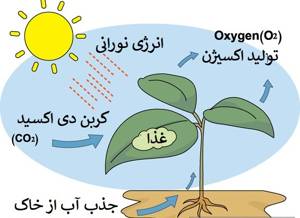
فعال ترین بافت فتوسنتزی در گیاهان عالی، مزوفیل برگ است. این سلول ها انرژی خورشید را توسط رنگدانه های سبز کلروفیل، جذب نموده و طی فرآیند فتوسنتز به ترکیبات آلی تبدیل می کنند. این ملکول های درشت، ذخیره کننده انرژی مورد نیاز برای برقراری زنجیره حیات هستند و در صورت حذف یا بروز هر گونه اختلال در این فرآیند، حیات روی زمین نیز مختل خواهد شد. همچنین جذب مقادیر عظیم دی اکسید کربن در اتمسفر و ترسیب آن در بافت های گیاهی یکی از مهمترین فرآیندهایی است که به حفظ تعادل اکولوژیکی کره زمین کمک میکند.
میزان تولیدات فتوسنتزی در شرایط مختلف محیطی و در گیاهان مختلف، متفاوت می باشد. در این راستا سه گروه گیاهی با مسیرهای فتوسنتزی متفاوت شناسایی شده اند:
گیاهان سه کربنه: فرآیند فتوسنتز در این گیاهان تابع مسیر کالوین-بنسن است و اولین ماده تولید شده که حاصل احیای کربن میباشد، ترکیبی سه کربنه به نام فسفوگلایکولیک اسید است.
گیاهان چهارکربنه: اولین ماده تولید شده در فرآیند فتوسنتزی این گیاهان که تابع مسیر هاچ و اسلک است، یک قند چهار کربنه می باشد.
گیاهان CAM: مسیر فتوسنتزی این گیاهان متفاوت از دو تیپ دیگر بوده و اولین ماده تولید شده در فرآیند فتوسنتزی آنها اسید 4 کربنه می باشد.
با توجه به اینکه نزدیک به 40 درصد از وزن خشک گیاهان طی فرآیند فتوسنتز و از کربن تشکیل شده است، لذا فتوسنتز لازمه رشد و بقای کلیه گیاهان در طول بخش عمده ای از چرخه حیات می باشد. از طرفی گیاهان برای رسیدن به حداکثر میزان فتوسنتز خود نیاز به مقادیر کافی آب و عناصر غذایی، نور مناسب و درجه حرارت مطلوب دارند.
در برخی شرایط نامطلوب محیطی میتوان گروهی از گیاهان سازگار شده با آن محیط را مشاهده نمود. این مسئله نشان می دهد که همه گیاهان حد مطلوب مشابهی ندارند. به عنوان مثال، اشکوب های زیرین جنگل با حداقل نور و بیابان های گرم و خشک با حداکثر نور خورشید و حداقل رطوبت از این گروه هستند. بنابراین اهمیت مسیرهای مختلف فتوسنتزی در این مناطق کاملا قابل درک است.
مسیر فتوسنتزی در گیاهان سه کربنه
در این گروه، احیای دی اکسید کربن تحت تاثیر آنزیم رابیسکو با پیش ماده ریبولوز بی فسفات انجام میشود. در این فرآیند ATP تولیده شده در واکنش فسفوریلاسیون نوری، برای تبدیل ماده ریبولوز مونو فسفات به RUBP مصرف شده و با احیای دی اکسید کربن اسید فسفوگلیسریک را به فسفو گلیسر آلدئید تبدیل می نمایند. در این مسیر ATP حاصل از واکنش های نوری احیا میشود.
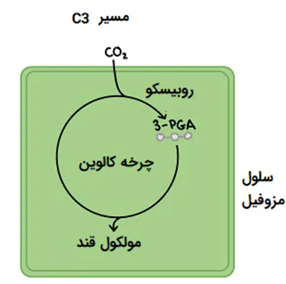
این شش مولکول ناپادار به سرعت به دوازده مولکول فسفو گلیسیریک اسید تبدیل میشود. این مولکول ها اولین ترکیب پایدار سه کربنه میباشد. در این مرحله دوازده مولکول فسفو گلیسریک اسید به دوازده مولکول قند آلدئیدی فسفات دار به نام فسفو گلیسر آلدئید تبدیل میشوند. این امر با صرف انرژی حاصل از مصرف ATP و NADPH صورت میگیرد.
سپس مولکول های حاصل از این فرایند به شش مولکول قند پنج کربنه ریبولوز بی فسفات و یک ماده قندی شش کربنه تبدیل می شوند که مولکول ریبولوز بی فسفات برای تداوم این سیکل در چرخه باقی مانده و مولکول شش کربنه میتواند به عنوان پیش ماده قندهای بزرگتر مانند نشاسته یا ترکیبات وابسته استفاده شود و یا در مسیرهای سنتز لیپیدها و اسیدهای آمینه مورد مصرف قرار گیرد.
مسیر فتوسنتزی در گیاهان چهار کربنه
با تغییر شرایط بوم نظام، افزایش غلظت اکسیژن و درجه حرارتی محیط، گیاهانی با سیستم های متنوع فتوسنتزی ظاهر شدند. در مناطق مختلف آمریکا بیش از 15000 گونه گیاهی با مسیر فتوسنتزی چهار کربنه که اغلب بوته ها و گونه های علفی می باشند، گزارش شده است.
در این گیاهان که 10 درصد از گونه های گیاهی را شامل میشوند، اولین مولکول حاصل از فرآیند کربوکسیلاسیون، ماده ای چهار کربنه به نام اسید اگزالواستیک است که ابتدا توسط هاچ و اسلک در سال 1966 کشف شد. در این گیاهان حلقه ای از سلول ها با دیواره ضخیم اطراف غلاف آوندی را احاطه میکند. در برخی از گونه ها دیواره غلاف آوندی در مجاورت این سلول ها ضخیم و یا چوب پنبه ای شده است.
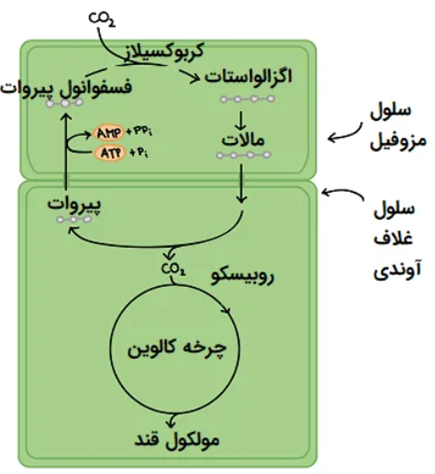
هرچند ممکن است نشت گاز اکسیژن از سلول های کلرانشیم به سلول های غلاف آوندی رخ دهد، اما وجود دیواره ضخیم چوبی شده برای کاهش انتشار اکسیژن الزامی به نظر نمی رسد. سلول های غلاف آوندی از طریق پلاسمودسماتا با سلول های مزوفیل مجاور که دارای دیواره نازک و فضاهای بین سلولی بزرگ می باشند، مرتبط شده اند.
به طور کلی یکی از تفاوت های گیاهان چهار کربنه با گیاهان سه کربنه، تفکیک مکان کربوکسیلاسیون و دکربوکسیلاسیون میباشد. آنزیم فعال فتوسنتزی در مرحله کربن گیری در سلول های مزوفیل، آنزیم فسفوانول پیروات کربوکسیلاز است که میل ترکیبی بسیار کمی با اکسیژن داشته و در نتیجه باعث کاهش بسیار زیاد در فرآیند تنفس نوری میشود. در این گروه از گیاهان واکنش اکسیژناسیون رابیسکو تنها در غلاف آومدی و به میزان بسیار جزیی مشاهده میشود.
گیاهان حدواسط سه کربنه-چهار کربنه
تا کنون بیش از 20 گونه گیاهی دارای مسیر حد واسط سه کربنه و چهار کربنه شناخته شده که از این میان میتوان به جنس های آلترناترا، پارتنیوم، موریکاندیا، پانیکوم و فلاوریا اشاره کرد. سرعت تنفس نوری در این گیاهان بسیار کمتر از گیاهان سه کربنه بوده و آناتومی کرانز تکامل نیافته است، اما آنزیم روبیسکو در سلول های مزوفیلی و غلاف آوندی مشابه هر دو گروه سه کربنه و چهار کربنه فعال است.
این گیاهان بر اساس نحوه باز زایی کربن به گروه تقسیم میشوند:
در گروه اول فعالیت آنزیم های فعال در مسیر گیاهان چهار کربنه حقیقی بسیار پایین بوده و فعالیت روبیسکو در سلولهای مزوفیل و غلاف آوندی بسیار زیاد است. بنابراین چرخه چهار کربنه در این گیاهان مشابه گونه های چهار کربنه حقیقی نیست. از میان جنس های ذکر شده میتوان به پانیکوم، آلترناترا و موریکاندیا اشاره کرد.
از گروه دوم دو گونه از نئوواکلما و فلاوریا شناسایی شده است. در این گیاهان آنزیم PEP کربوکسیلاز فعال بوده و فرآیند تغلیظ دی اکسید کربن مشاهده میشود، ولی میزان عملکرد کوانتومی این گیاهان کمتر از گیاهان سه کربنه و چهار کربنه است. برخی گونه های گیاهی مانند الوکاریس و جنس آتریپلکس برای تثبیت کربن در اندام های مختلف از هر دو مسیر سه کربنه و چهار کربنه استفاده می کنند.
مسیر فتوسنتزی در گیاهان CAM
گیاهان این گروه برای تثبیت دی اکسید کربن از مسیر متفاوتی نسبت به دو گروه سه کربنه و چهار کربنه استفاده مینمایند، بدین ترتیب که شب هنگام، نسبت به عمل کربن گیری اقدام کرده و اولین ماده تولید شده در فرآیند تثبیت کربن یک اسید چهار کربنه مانند اسید مالیک می باشد. مهمترین گیاهان این گروه از چهار خانواده فرفیون، ارکیده، آناناس و کاکتوس می باشند.
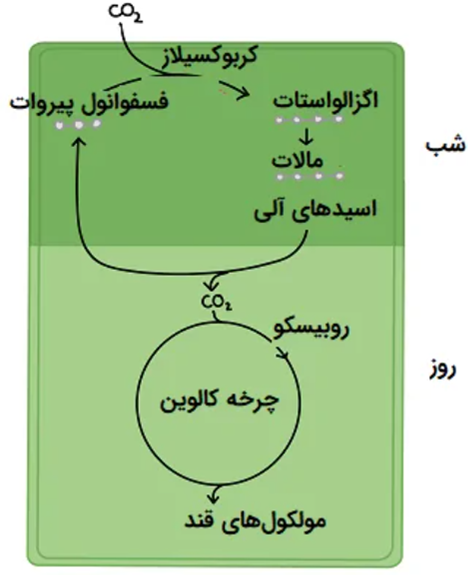
آناناس زراعی نیز در این گروه قرار دارد. گیاهان کراسوله بسته به شرایط محیطی به دو گروه CAM اختیاری و CAM اجباری تقسیم میشوند. در گروه CAM اختیاری گیاه در صورت مطلوب بودن شرایط محیطی میتواند از مسیر فتوسنتزی سه کربنه استفاده نماید. برخی از گیاهان نیز با تغییر شرایط محیطی، تابع مسیر فتوسنتزی چهار کربنه میشوند. ولی در گروه گیاهان CAM اجباری، با تغییر شرایط محیطی تغییری در مسیر فتوسنتزی بوجود نمی آید.
به طور کلی ویژگی های گیاهان کراسوله عبارت است از:
1-باز شدن شبانه روزنه ها برای تبادل گازی با محیط
2-تغییرات غلظت اسیدهای آلی(غالبا اسید مالیک) تولید شده طی فرایند تثبیت کربن در طول شبانه روز
3-تغییرات غلظت قند و نشاسته بر خلاف نوسانات اسید مالیک
4-فعالیت بالای آنزیم PHP-کربوکسیلاز در شب
5-وجود واکوئل های بزرگ در سلول های کلرانشیم و محتوای متفاوت آب در اندام های مختلف(برگ، ساقه و طوقه) بسته به جنس گیاه
گیاهان کراسوله غالبا در مناطقی رشد می کنند که از نظر رطوبت و درجه حرارت شرایط نامساعدی دارند. بنابراین تولیدات فتوسنتزی در این گیاهان نسبت به دو گروه سه کربنه و چهار کربنه کمتر است. بر این اساس اگر این گروه از گیاهان در شرایط مناسب آب و هوایی پرورش داده شوند، قابلیت تولید بالاتری خواهند داشت. به عنوان مثال، آناناس در شرایط رطوبتی مناسب، عملکرد بیش از گیاهان چهار کربنه دارد. در این بین تنها یک جنس از گیاهان چهار کربنه به نام خرفه نیز وجو دارد که در شرایط خاص از مسیر CAM استفاده میکند.
گیاهان کراسوله در شب به دلیل کاهش درجه حرارت محیط، تعرق کمتری داشته و اقدام به باز نمودن روزنه ها میکنند. بنابراین با جذب دی اکسید کربن، اسید مالیک تولید شده در واکوئل ها ذخیره می شود. با آغاز روز، اسید مالیک ذخیره شده طی شب به مواد آلی مورد نیاز فتوسنتز و تولید قند تبدیل شده و به کلروپلاست منتقل می شود.
بنابراین با گذشت زمان در طول روز، از غلظت مالات (اسید آلی) کاسته خواهد شد، چرا که به دلیل کاهش ذخیره مالات واکوئلی و تبدیل آن به قند، غلظت مالات در نیمه شب به حداقل خود میرسد. بنابراین در صورتی که گیاه در تاریکی قرار گیرد، از کربوهیدرات های ذخیره شده در طی روز استفاده میکند، به طوری که با افزایش مدت زمان تاریکی، مقدار نشاسته ذخیره شده هر دو گیاه سه کربنه و CAM به حداقل میرسد.
بر این اساس گیاهان CAM تنها شب هنگام اقدام به جذب دی اکسید کربن نموده و انرژی مورد نیاز خود را نیز از سوختن مواد نشاسته ای دریافت می نمایند، ولی گیاهان سه کربنه طی شب تنها به فرآیندهای تاریکی فتوسنتز و تولید مواد قندی پرداخته و طی فرآیندهای مستقل از نور و هم زمان با استفاده از مواد نشاسته ای ذخیره ای به تولید این مواد نیز می پردازند. بنابراین میزان ذخیره نشاسته در گیاهان سه کربنه نسبت به گیاهان CAM کمتر است، ولی سرعت کاهش ذخیره نشاسته در گیاهان CAM بیشتر است.